Chapter 12
Theory of Consciousness
Mindesign
In Chapter 7, we found that there is one belief we can hold with certainty, the belief that I do exist. Conscious minds soon learn that while “I am” as a thinker, there also exists an “other” that is affecting my thoughts. My consciousness discovers that it is matter bound. Externally caused motions of the material body to which I am bound play a role in the thoughts of my non-physical consciousness.
My consciousness learns too that it has the reciprocal capability of governing the motions of the matter to which it is bound. The I that thinks is the I of a matter-bound consciousness. The mind and the body it controls form a conscious body of matter, the minderelle. Our entire conscious experience is bound to only a handful of atoms.
In the section “The Brain’s Command Center,” from Chapter 3, where we were discussing the primary minderelle of humans, we found that “The consciousness of this one molecule at the command center will make all the decisions, feel all the emotions, see all the sights and hear all the sounds; it will experience hunger and thirst, it will taste, smell, and feel the effects of any drugs and alcohol, and in every other way, this one molecule will experience the full life of its human body.” For this to be true, all the sensory data collected by the body must be funneled into this one primary amino acid continuously. Simultaneously, we are also in command of our human body, so this same amino acid must be sending out a continuous stream of commands for all the actions to be taken by the body.
Our finding that consciousness is bound to so few atoms raises serious questions. How could the many functional requirements of the human primary be handled by so few atoms? Would it even be possible for so few atoms to handle so much incoming sensory data while simultaneously generating all the outgoing command data needed to direct the actions of the entire human body?
Aside from the capabilities needed to act as the primary of a human, there are many other problems that still have to be addressed.
Geometry locks were established in Chapter 5 as the method conscious minds will use to change material outcomes. So why do we not have any experience of locking or unlocking the distance between two atoms? Minderelles experience constant collisions with surrounding matter, so can geometry locks be held during collisions without violating the conservation laws? How is it we can see a vision when our consciousness belongs to so few atoms?
These and many other questions remain. In short, we must show that it is possible for consciousness to provide all the rich life experiences we are familiar with even while restricted to so few atoms.
This chapter is dedicated to “mindesign,” the subject dealing with the structure and operational capabilities of consciousness.
The Binary to Sensory Conversion
Earlier, we found that information is transferred between minderelles in the form of binary 1’s and 0’s. But how do we get from the binary 1’s and 0’s entering our minderelle to an extreme hunger experience after going days without food? When a large rock falls on our leg, how do we make the transition from the incoming binary data to “feeling” pain? We do not learn there is a pencil on the table by analyzing reams of incoming 1’s and 0’s. Instead, we simply see a pencil on a table.
If all sensory data enters our minderelle in the form of 1’s and 0’s, then how does our consciousness magically transform these raw 1’s and 0’s into visions, sounds, taste, pain, hunger, and other high-level sensory experiences?
The fact that these high-level sensory experiences are so far beyond the format of the incoming raw data reveals that consciousness is capable of processing raw data and converting it into the high-level conscious experiences we are familiar with. To make this conversion, consciousness itself must have structure and high-level operational capabilities.
Methods of This Chapter
Is it reasonable to think that all the interactions between our consciousness and our surroundings take place through the few atoms of a single minderelle? The answer to this question will depend on whether we can find a mindesign solution that makes this claim reasonable, so our goal in this chapter is to find a mindesign solution that makes reasonable all the claims we have made in this free will paradigm. So how will we judge whether a mindesign solution is reasonable?
Because we found that god loves us, we will first accept that god designed consciousness to have all the operational capabilities needed for us to survive and thrive in the universe. Next, we will find that the design of consciousness required solutions to the same problems that humans would later have to solve, which range from creating functional societies to maximizing the usefulness of computers and electronics.
So to proceed from here, we will accept that like minds using like reason solve like problems alike.
If a solution to a problem was reasonable for humans, then we will accept that the same solution to the same problem would have been reasonable for god. Furthermore, if humans took the time and trouble to create capabilities to better the lives of other humans, then we will accept that a loving god would have taken the time and trouble to create the corresponding capabilities in mindesign.
In other words, we will accept that human design mimics mindesign. To figure out what god did with mindesign, we will start by asking ourselves, what would humans do? This makes sense because we have accepted that we were made in the image and likeness of god, and “like minds using like reason solve like problems alike.” This approach will be our guide. We will consider our societies, our computers, and our electronics to be recreations of the structure and operational capabilities of consciousness. That which we have done, so did a like-minded god also do.
The Television and the Mindshell
There are two major components to a complete television system; there is the television itself, and there is the conscious viewer. Of these two components, it is the television that serves the conscious viewer, not the other way around. So what function, exactly, does the television offer that makes it useful to the viewer?
We start by noting that all sound and video data enters a television as a continuous stream of 1’s and 0’s, so incoming television data has the same format as the data entering a minderelle.
The role of the television is to convert these incoming 1’s and 0’s into sound and video images. But instead of converting the incoming 1’s and 0’s into images, why not replace the entire television with a simple light that flashes on and off with the incoming 1’s and 0’s? Why not just let the conscious viewer extract the video and sound information from the flashing light and skip all the electronics? After all, the flashing light would contain all the same information as the television images and sound, only in a different format.
The answer may seem obvious, but it is important. Even if the conscious viewer could keep up with the high rate of flashes, and even if the viewer had the capability of extracting the encoded sound and video information from the flashing light, free-thinking minds are easily distracted. The 1’s and 0’s are arriving at 100 million per second, and every time the viewer loses track of even one flash, large swaths of information would be lost. Free-thinking minds simply do not have the attention span and focus needed to extract meaningful data from raw streams of 1’s and 0’s.
God faced this same dilemma while working on the mindesign problem. To comply with the conservation laws, god was limited to using geometry locks for communication between minderelles. Information transmitted via geometry locks are also in the form of 1’s and 0’s because geometry locks only have two states, locked and unlocked.
Therefore, the data entering the primary minderelle was the same as the raw data entering modern televisions, both were in the form of binary 1’s and 0’s. But if an easily distracted mind could not make sense of the 1’s and 0’s entering the television, neither could this same mind make sense of the 1’s and 0’s arriving from the brain as visual and audio data.
Let us now reconsider the two components of the complete television system. The viewer is the conscious component of the television system, while the television itself is the fully determined component. That is, to get useful information from streams of 1’s and 0’s, the conscious viewer needs the fully determined television component to convert the incoming 1’s and 0’s into the easily digestible formats of video and sound.
Similarly, god also needed a fully determined component of consciousness that would perform the same function as the fully determined television, a mind component that could deterministically convert incoming 1’s and 0’s into easily digestible formats, or sensory fields. Just as a complete television system has two components, the television and the viewer, so would consciousness also require two analogous components.
We will call the fully determined component of consciousness the mindshell, and we will call the free-thinking component of the mind the mindcore. The mindcore and the mindshell are the two major components of consciousness, so both are massless and momentum free.
Figuratively, the mindcore is at the center while the mindshell envelops the mindcore. Thus, all incoming data will first be processed by the outer mindshell before it passes through to the mindcore at the center. For example, incoming visual data formatted as 1’s and 0’s will be processed by the mindshell and then directed to the visual sensory field of the mindcore in a format the free-thinking mindcore can easily digest. Similarly, outgoing command data generated by the mindcore will be processed by the mindshell so it will be properly formatted for export to the brain.
The Mindshell:
The component of consciousness that carries out all the functions of consciousness that are fully determined and programmable.
Meanwhile, our thoughts and sensational experiences are the thoughts and sensational experiences of the mindcore.
The Mindcore:
The free-thinking component of consciousness that experiences life, makes choices, and is the personality of the minderelle.
We will soon find that the mindshell is needed to carry out many other fully determined processes not yet considered.
Memory
To establish the memory capabilities of the minderelle, we will step briefly away from human considerations and consider operational proteins.
Proteins are made up of hundreds or thousands of minderelles, each of which is governed by an independent mind. Because each minderelle can only govern the motions of its own amino acid, the functional motions of the full protein will require that all the minderelles of the protein work together to coordinate their actions.
However, the individuals within the protein could not coordinate their actions unless each were to know its own current reality. That is, for a protein to function effectively, each mind in the protein chain would first have to know that it belongs to a group of like individuals that form a larger body, the protein, and that this larger body has a function. Each individual must also know that “I” play an indispensable role in helping this larger body carry out its function.
There is one necessary condition that must be met before a mind can grasp its own current reality and learn to work as part of a team to help make “my” protein functional. That necessary condition is memory. If the minds in the protein chain were to have no memory, then the passing of each moment in time would be the same as having a chain of newly conscious beings coming into existence continuously.
This equivalent of a long chain of individuals rebooting their conscious existence at every moment would make it impossible for them to get a grasp of their current reality. If each minderelle did not have a memory to tie the present moment to past moments, there could be no understanding, no learning, no cooperation and no common function. Without a memory, it would be impossible for the minderelles of a protein to coordinate their actions for the good of the whole.
However, the minderelle does not have sufficient material complexity within which fully functional memories could be encoded. For this reason, if this free will paradigm is correct, memory must be an inherent part of consciousness. Just as humans add memory capabilities to computers and electronics, so would god have added memory as an operational capability of consciousness.
This free will paradigm now accepts that memory is an inherent operational capability of the conscious mind.
Sensory Fields
All information enters a minderelle as 1’s and 0’s, but we have no awareness of any incoming binary data stream in this raw format. Instead, we see a pencil on a table, we taste a pineapple, hear a voice, experience hunger, and we smell the flowers – we have sensory experiences. But how can we “see” a pencil when our consciousness belongs to only a handful of atoms?
The physical body of the minderelle does not have the complexity that would be needed to create any of the complex sensory experiences we are familiar with. Therefore, the complexity required for our sensory experiences must reside on the side of the massless and momentum-free mind. In the previous section, we found that memory had to be an operational capability of the mind. We now find that consciousness has built-in sensory field capabilities as well, the most familiar of which are vision, hearing, taste, smell, and touch.
Just as the television deterministically converts incoming 1’s and 0’s into visual images that are displayed on a screen, so too will the mindshell deterministically convert incoming visual data in the form of 1’s and 0’s into visual images and display them in the visual sensory field. The mindshell will similarly use audio data arriving as 1’s and 0’s to populate the audio sensory field, and so on with all the incoming sensory field data.
We may also have other sensory field capabilities we are not aware of. For example, the primary of a worm will have a visual sensory field just as we do because all like bodies must function alike. But worms evolved in an environment where the visual sensory field was apparently not put to use.
Similarly, we may also have sensory fields that are not in use due to the environment in which we evolved. For example, conscious minds may have a sensory field that would be ideally suited for presenting the mindcore with information about our surrounding magnetic field. The human engineering minderelles may have chosen to leave this sensory field unused because the biological cost of the needed infrastructure would be too high for so little gain in survival. However, this might not be the case for some migratory species.
This free will paradigm now accepts that the mind has operational capabilities that include at a minimum the memory and the sensory fields.
The Constants of Consciousness
The mindshell of consciousness is the same for everyone and is fully determined. The fully determined mindshell establishes the universal constants and handles the operational requirements of the mind/matter interface for all thinking minds in the same and knowable way.
For example, the mindshell sets the speed of thought progression so it is the same for all conscious minds for any given set of conditions. It is also the mindshell that makes both the conscious and the sensory experiences the same for all conscious minds. If there were no constants of consciousness, one mindshell could translate incoming data as a beautiful visual scene while a second could translate the same incoming data as pain.
Without a universal and fully determined mindshell, the experience of emotions we observe in others could be different from our own experience of emotions, but this would make consciousness unknowable. Having the mindshell regulate the constants of consciousness does not have any effect on who the thinker is, rather, it sets the stage from which all conscious individuals throughout time and space experience life. It creates a common operating environment from within which all thinking minds live and function.
The Mindshell Imposes Geometry Locks
In Chapter 5, we found that minds act on the matter of their minderelles by imposing geometry locks. This means that the commands needed to put a humanon body in motion must exit the primary minderelle in the form of geometry locks. However, when we make a decision to do something, we have no awareness of atoms and bonds, geometry locks, or of anything at all happening at the atomic level. We make the decision to walk and walking happens. We make decisions, and then somehow, that is what the body does. So how do we get from the high-level decisions made by our thought processes to the geometry locks that are required at the scale of our minderelle?
Let’s look at what happens inside today’s computers. Today’s programmers use high-level languages that resemble human languages. But before the computer can execute these high-level programs, the program must first be converted, or compiled, into low-level machine language that computers can work with. At the machine language level, all instructions are in the form of binary 1’s and 0’s. This language conversion that takes place inside computers is a deterministic process carried out automatically, a process today’s programmers are not involved with.
And so it is with us when we make decisions that have to be exported to the brain using geometry locks. Recall that it is the role of the mindshell to carry out all the functions of consciousness that are fully determined and programmable.
Thus, it is the mindshell that handles the equivalent of the compiler code in a computer. Our conscious mindcore thinks freely and makes decisions. The mindshell then deterministically converts these high-level decisions into the low-level machine language that can be exported through geometry locks.
The Brain/Primary Interface
We have now introduced the structure of consciousness at its most basic level, which consists of a determined component and a free-thinking component. We have also introduced some of the operational capabilities of consciousness, including memory and the sensory fields. We now need to cover this material in detail, and we also need to tie up many loose ends from earlier chapters.
Because our conscious experience is specifically human, we will continue to focus on the human primary and the human brain for much of the upcoming material. This upcoming material will also require a basic understanding of how the human primary interfaces with the brain. Before we begin our discussion on the interface between the brain and the human primary in the following sections, it is important to emphasize that this brain interface evolved over time and is part of the brain, it is not a part of mindesign.
Mindesign deals with the structure and operational capabilities of consciousness and is entirely the work of god. The brain/primary interface, on the other hand, deals with the portion of the brain adjacent to the human primary and is the work of minderelles. The brain/primary interface is where information from the brain is consolidated for input into the primary, and it is also where commands from the primary are divided out into major functional groups for distribution to the various parts of the brain.
Because this interface is the result of billions of years of evolution, some specifics of the brain/primary interface will be unique to each species. However, the brain/primary interface must be operationally compatible with the already existing mindesign of the primary, so the interface for each brained species will be operationally the same.
Time Division Multiplexing
Earlier we found that each minderelle only has three communication links, which were the two shared bonds and the one hydrogen bond pair. However, more than three types of data are continuously entering the primary minderelle – vision, audio, taste, smell, memory, appetites, and so on. Furthermore, this same primary must also export command data continuously.
How can so many types of data be entering and exiting the primary minderelle when the minderelle only has three communication links? And if all the incoming data must enter the primary as 1’s and 0’s, how will the primary know which 1’s and 0’s correspond to which data type?
The minderelle has fewer communication links than incoming data types, so to be functional, the mindshell will have to have a timer cycle that continuously cycles through all the operational capabilities of consciousness. With a cyclical timer, each operational capability of the mind can now be given its own input time slot.
The brain had to evolve in a way that ensured each data type would arrive at the primary during its corresponding time slot. That is, the brain must supply visual data to the primary during the visual time slot, audio data must arrive at the primary input during the audio portion of the timer cycle, and so on.
Each type of sensory data from the body is processed in a different part of the brain, so the brain will have to combine all this data sequentially in the order of the corresponding time slots so the combined data can then be fed into the primary as a single, continuous data stream. The brain will combine all this data into one data stream using the same solution used by humans in communication electronics, it will use a multiplexer.
In electronics, there are different types of multiplexers, but only one is suited for our purposes here. When independent data streams are each given their own time slot to a common transmission line, the process is known as time-division multiplexing. The brain’s multiplexer will likewise give each type of sensory data its own time slot so that each will be supplied to the primary during its corresponding time slot. This data will enter the primary through the shared bond dedicated to incoming data. The process is outlined in Figure 23.
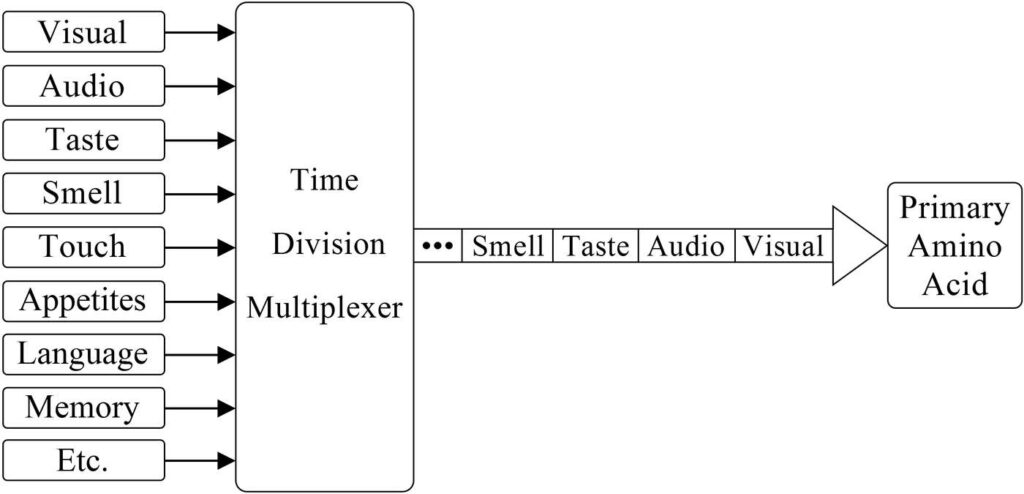
Figure 23. The multiplexer combines sensory and other data streams onto a single line for entry into the primary. The many independent data streams are arriving from the brain at the left side of the multiplexer shown above. The multiplexer is giving each data stream its own time slot for accessing the common line to the primary. The multiplexer is a physical part of the brain, it is not a part of consciousness.
In each complete cycle of the multiplexer, one complete visual image will pass through to the primary, then one time span of audio data, then one capture of taste data, and so on. When the timer cycle has completed a full rotation through all the data types, it will then begin a new cycle in a rotation that repeats. Time-division multiplexing makes it possible for humanon primaries to be continuously updated with the newest information coming in from all parts of the humanon body.
An inverse process will be taking place with the shared bond on the output side of the primary. Commands for directing the actions of the body are issued by the mindcore of the primary. The mindshell of the primary then converts the high-level commands from the mindcore into binary 1’s and 0’s so they can be exported using geometry locks through the shared bond dedicated to outgoing command data. This single stream of complex command data from the primary will then be fed into a demultiplexer that will break the commands out and distribute them to the various parts of the brain.
Conscious Continuity
General readers not familiar with timer cycles in electronics may find it hard to imagine that our consciousness could be divided up into discrete, repeating time slots. However, timer cycles in computers and electronics are ubiquitous. The key is to make the complete timer cycle so fast that the conscious mind cannot tell that any cycling is taking place.
For example, when we watch videos on our phone, the phone display is stepping through a series of still images at some timer cycle rate (the refresh rate), and this refresh rate is so fast that the conscious mind does not notice that the video is made up of still images.
Just as human engineers have incorporated timer cycles into virtually every aspect of computers and electronics, so would god have made use of the same highly functional solution. Again, the guiding assumption of this chapter is that like minds using like reason solve like problems alike. Therefore, we accept that the same timer cycle solutions that are so commonly employed by humans would have been just as reasonable for god.
The Primary’s Hourglass Beta Sheet
In Figure 23 above, the time-division multiplexer is shown as a simple box, but what is inside this box? Figure 24 illustrates a simplified, but fully functional design for both the multiplexer and the demultiplexer. Both are broken out into individual minderelles, shown as the light blue outlines. The hydrogen bonds are shown as the vertical dashed line.
Figure 24 shows how all brain data can pass through the single amino acid of a humanon primary. In this figure, the primary is the red minderelle located at the center of the hourglass shaped beta sheet. The minderelles on the left side of the figure act as the multiplexer by combining all the data headed to the primary onto a single data line, with each input being given its own time slot. Notice the indispensable role of the hydrogen bonds in the information routing process.
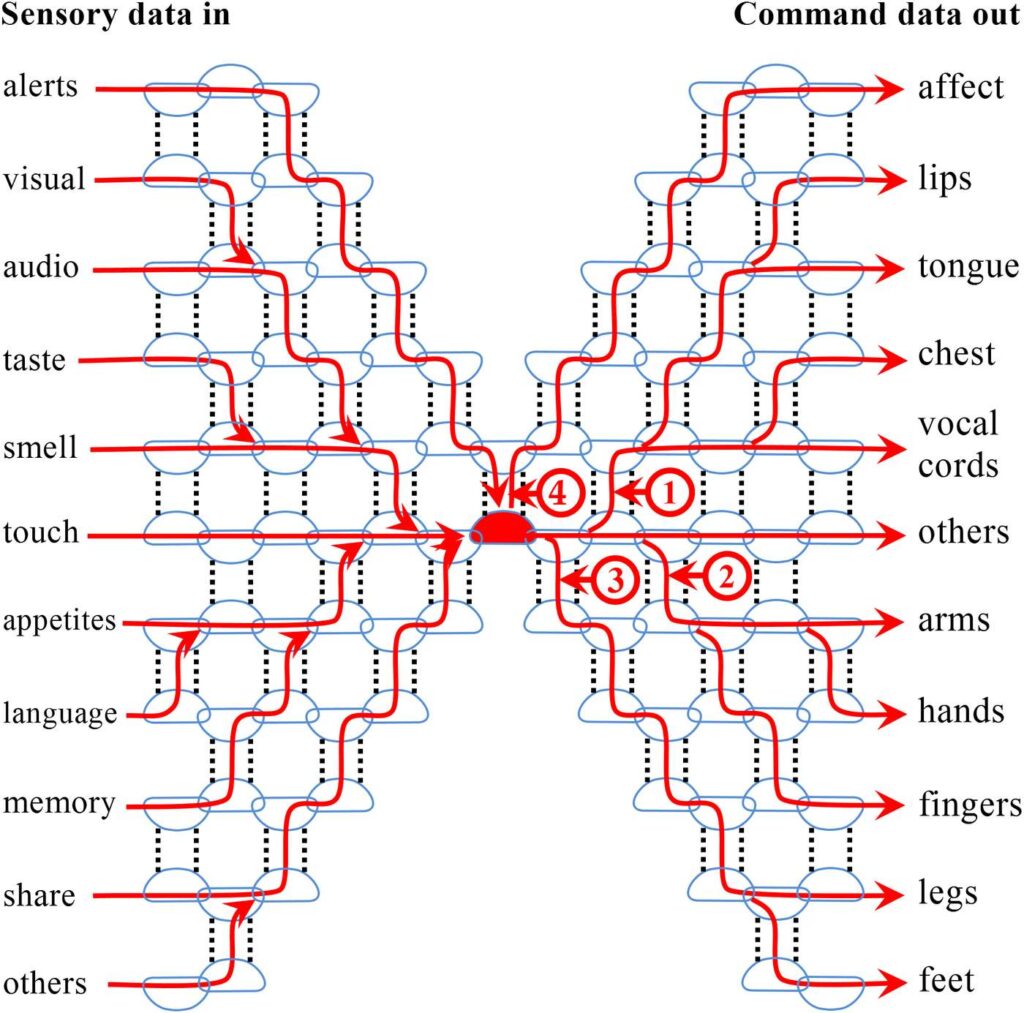
Figure 24. The humanon primary is the red minderelle at the center, other minderelles are shown only as light blue outlines. The many data streams are shown with red lines. The multiplexer is to the left of the primary, the demultiplexer is to the right. Notice the hourglass shape of the primary’s beta sheet, which makes it possible to funnel all brain data through the one minderelle that is the primary. Also notice the indispensable role played by the hydrogen bonds. The time-dependent switching is not shown.
The data arriving at the multiplexer inputs on the left has already been fully processed by the brain. While the figure shows all the data being funneled onto one common pathway, what is not shown is the time-dependent switching that allows only one data stream to access the primary at a time.
The minderelles on the right side of the figure act as a demultiplexer, which divides the command data from the primary into different functional groups that will then be fed into the appropriate areas of the brain for processing. For illustration purposes, we speculate on several major muscle groups for humans in the figure: (1) the voice functional group, (2) the upper extremities, and (3) the lower extremities. As with the input side, a time slot is allotted to each functional group.
The process is no different than the way demultiplexers work in electronic circuits created by humans.
The number 4 in Figure 24 identifies the hydrogen bonds of the primary. The hydrogen bonds of the primary may offer contextual information for both incoming data and outgoing data.
For example, if the visual cortex within the brain detects the face of a loved one while processing the visual data, the mindcore of the primary will see the face of the loved one in the visual images that arrive from the multiplexer through the incoming shared bond. Meanwhile, the hydrogen bonds of the primary will provide the name and the nature of the relationship, information drawn from the memory storage areas of the brain.
The mindshell of the primary would be tasked with deterministically joining the shared bond input and the hydrogen bond input for final presentation to the mindcore.
The Timer Cycle of Consciousness
We found that time is allotted to each of the operational capabilities of consciousness in a repeating cycle that is governed by the mindshell. All of this exists within the momentum-free portion of the minderelle, so it is part of mindesign. Therefore, the finding that the mindshell of consciousness has a repeating timer cycle will apply to all conscious minds and has since the beginning of time.
Do Red Roses Exist?
Do red roses exist?
This question is not asking whether the color red exists. We see in color, we have labeled one of those colors red, and therefore we accept that the color red exists.
Neither is this question asking if roses exist. Many species of flowering plants exist, we have labeled one of those species the rose, and therefore we accept that roses exist.
So we accept that the color red exists, and we accept that roses exist, but do red roses exist? Over the next several pages, we will argue that red roses do not exist.
Understanding that red roses do not exist is important for a number of reasons. It will help us understand the sensory fields of consciousness more clearly, and it will also help us better understand the brain’s capabilities when it comes to representing the body’s surroundings to the primary.
Swapping Multiplexer Inputs
As the brain evolved, each species was able to tailor its own version of the brain to most benefit its own survival. This has far reaching consequences. In Figure 23, we showed a time-division multiplexer that makes it possible for all sensory data to enter into a single amino acid, the primary.
But what would happen if a species chose to swap the inputs to the multiplexer? What if a species chose to put audio data into the visual data time slot, or visual data into the audio data time slot? Could the mind “see” sound, or “hear” a vision? And could a species benefit by switching the input lines?
We will now consider the benefits of switching the input lines to the multiplexer.
Sonar mapping
Scientists use sonar to map the ocean floors by directing bursts of sound towards the ocean bottom. The echoes of these sound bursts are detected by directional microphones which collect the data to create maps of the ocean floor. No individual component of this sonar mapping process is beyond the known capabilities of currently existing biological humanons here on Earth.
Therefore, it is reasonable to think that somewhere in the universe, humanons exist that are biologically capable of sonar mapping. Considering how beneficial this sonar mapping capability would be for bats living in dark caves, and to make the point of this section, we will make the assumption that somewhere on Earth, there are bats that have developed a sonar mapping capability.
Our mapping bats will have many directionally sensitive microphones distributed throughout the external structures of their ears, perhaps in the form of cilia tuned to vibrate directionally at the bat’s call frequency, or in the form of microsized sound pressure detectors located under the skin of the curved external ear structures.
The ear structures of our mapping bats will be curved to bring about the amplitude and timing variations in the reflected sound that would be needed for direction and distance information to be extracted. As for the sound used for echo mapping, scientists use high-frequency sound bursts, and as luck would have it, real bats already create similar sound bursts at similar frequencies.
The data from the array of microphones located throughout the internal surface of the two ears will be carried by nerves directly from the ear structures to the brain. This separate nerve network means that the mapping capability of our mapping bats will operate independently of the bat’s normal hearing capability.
So we now have special bats that generate mapping worthy sound bursts, they have directionally sensitive microphones for collecting echo data, and they have brains for data analysis. Therefore, our hypothetical bats are now cave mapping capable.
The question we now wish to address is this: What does the brain of our mapping bat do with its sonar generated map of the cave?
The answer for our bats is likely the same answer found by human scientists. Like minds using like reason solve like problems alike. Scientists prepare their sonar maps of the ocean bottom for the visual sensory field. That is, scientists format their mapping data that was created from sound for presentation to our visual sensory field. This seems inconsistent with our intuitive understanding of the sensory fields.
Normally we think that data from light should go to our visual sensory field, data from sound should go to our audio sensory field, data collected from independent molecules suspended in the fluid surroundings of our bodies should go to the olfactory sensory field, and the sampling data collected from the solids we eat should go to the taste sensory field. And yet, scientists prepare their mapping data that came from sound for presentation to our visual sensory field.
And so will the brains of our mapping bats also direct their sonar mapping data to the visual sensory field of the primary, and for the same reason. The visual sensory field is best suited for handling two dimensional maps, and even more so considering that the visual sensory field of consciousness includes the added dimension of color.
This means that the brains of our mapping bats will be able to select between two sources for loading the visual sensory field of the bat’s primary. One source for the visual sensory field will be the eyes, the other source will be the sonar data. The mapping bat’s brain will switch to the eyes whenever there is sufficient light because light would offer superior imaging. When light levels are too low, our mapping bat’s brain will switch to sonar data.
Using Memory for Sonar Data Expansion
Scientists store their sonar created maps of the ocean floor in computer memory. Eventually, scientists will have a complete and detailed map of the ocean floor stored in computer memory. If these scientists are now presented with limited mapping data of a small, localized site from the ocean floor, they will find the location of this site by searching for a match within the already existing global map.
Biological brains also have memory and they also have the processing capabilities needed to carry out the same search and match function. Brains do this all the time with faces. Thus, when our mapping bats are flying inside their already mapped caves, their brains will match the limited data from the bat’s live sonar to the fully mapped cave data from memory. This will make it possible for the bat’s brain to continuously display the full cave in the bat’s visual sensory field during flight, even though the bat’s sound bursts are narrowly focused.
The bat’s brain needs only enough live sonar information from the cave walls to keep the bat’s complete vision of the surrounding cave always current. Therefore, the brain’s search and match function will make it possible for the bat to keep its sound bursts narrowly focused, and this will improve the bat’s ability to target prey, but without compromising the bat’s ability to navigate within the confines of the cave.
Colors by Assignment
Scientists present their sonar mapping results in color. Interestingly though, there is no natural correspondence between colors and the sound data collected, so scientists assign colors in a way that promotes a natural understanding of their data. Colors may be assigned according to the height of the ocean floor’s features, or color-coded according to the type of vegetation or rock matter at each location on the floor. What is important here is that colors are assigned by the scientists so they will most intuitively represent the findings the scientists are focused on.
So how will bat brains present the cave image to the bat primary?
We think of bat caves as dark places, so it is hard for us to imagine that our mapping bats could see their caves as brightly lit and in full color. However, surely they would. Consciousness offers the whole rainbow of colors for viewing the visual sensory field. We know this by personal experience, we see the visual sensory field in color.
So once the bat has fully mapped the cave, the bat’s brain can process the map so it will display to the primary in whatever manner best aids navigation. For a fast flying bat confined to the small space of a cave, we would expect the bat’s brain to display a brightly lit cave with colors that will be most helpful in avoiding collisions with the cave walls.
For example, the bat may see features farthest away as violet, with the other colors then applied in the order of the rainbow according to distance. This means the closest objects would appear red. These same colors could also be assigned in reverse, colors could be assigned to represent the relative velocity of objects rather than their distance from the bat, and so on.
If a human were now to put a red rose in the cave, what would be the color of this rose to the bat?
If the rose’s color is assigned by the distance between the rose and the bat, then the rose’s color will change according to the bat’s distance from the rose. If the rose’s color depends on the relative speed between the bat and the rose, then the rose’s color will change according to the bat’s speed. The color of the rose to the bat will depend on whatever color assignment scheme was selected by the bat’s brain during evolution.
But the same thing can be said about the way human brains process the photons that come from the rose. That is, there is nothing about the rose itself that makes it red. We see it as red only because that is how the engineers of our brain chose their color assignment scheme. The color assignment scheme that was right for us may not be right for humanons that evolve in other environments or with other needs. In other words, color does not exist in the object, it exists within the consciousness of the mind.
In nature, there are simply photons and sound waves with energies spread across a continuous spectrum. There is nothing that creates a natural correspondence between the photon spectrum and the colors we can perceive in our visual sensory field. The visual sensory field of consciousness has color capabilities, and the brain of each species will manipulate all visual sensory data before it enters the visual time slot of the primary so that colors will be displayed in a manner that best suites the species.
This means that the color of the rose is not in the rose, but in the consciousness of the mind, and it will be displayed according to whatever color assignment scheme was chosen during evolution. The rose is in your hand, but the red is in your head.
Sensory Field Crossover
More generally, a minderelle designed brain can analyze data from any source and then direct this data to the multiplexer input line of its choice. When the nose of a human detects chemicals in the air, the findings are presented to the olfactory sensory field. In other species, these findings could be sent to another sensory field, or to multiple sensory fields.
For example, when dangerous chemicals are discovered in the surrounding air or water, the brains of some species may send a signal to the audio sensory field to sound an alarm with a pitch assigned to that specific chemical.
If the eyes of a humanon evolved to have spectroscopic capabilities that could detect chemicals in the air, this visual data could be directed to the olfactory sensory field as a smell that corresponds to the chemical, or again, it could be directed to the audio sensory field in the form of a background frequency corresponding to the chemical, or it could be directed to the visual sensory field as a colored ring around the outer visual field.
All of this is possible because the mindshell of the primary will process the data arriving from the multiplexer in always the same, determined way. This makes it possible for the brain of a species to manipulate the data arriving at the inputs of the multiplexer in a manner that will achieve the desired sensory effects.
That is, how sensory data is perceived in each sensory field of the primary depends on how the brain chooses to manipulate the inputs of the multiplexer. With this being the case, we should keep in mind that when we discover humanons that evolved in a vastly different environment, they may have ways of informing themselves of their surroundings that we do not have.
The Local Communication Limitation
With our current understanding of consciousness, each minderelle in a protein is only able to communicate with its adjacent neighbors. However, proteins carry out complex functions as a whole, and proteins are hundreds or thousands of amino acids long. So how does the primary of a protein command the actions of these hundreds or thousands of individuals so that the protein can carry out its intended function as a whole?
We will assign the primary of a protein position 0. Now consider a single command message sent from the primary to the amino acid at position 300. The primary starts by sending the message to the minderelle at position 300, but before the message can get to minderelle 300, it must first pass through the minderelle at position 1. So mindcore 1 gets the message and understands it is intended for 300. Mindcore 1 now repeats the message and sends it on to minderelle 2. The conscious mindcore of minderelle 2 receives the message and realizes it is intended for 300, so it repeats the message as best it can and forwards it on to minderelle 3, and so on, until the message finally gets to amino acid 300.
But for the protein to do its work, the primary will be sending command messages to all the minderelles in the protein. Simultaneously, all the minderelles of the protein will be sending sensory updates and feedback messages in the other direction to the primary.
So we now have command messages leaving the primary continuously for every minderelle in the protein, and feedback messages from all these minderelles heading in the other direction to the primary. This means that the mindcore of every amino acid has to carry out its own work assignments while also passing along a never ending stream of messages in both directions continuously.
This reminds us of the telephone game where a starting player whispers a sentence to the next player, who in turn whispers it to the next player, and so on until it reaches the last player. Anyone who has played this game knows that when messages are repeated many times, the results can be comical.
But now imagine the players trying to pass along a stream of messages in both directions in rapid fire succession continuously. The results would be unintelligible. And the results would be no different in proteins. We are them and they are us.
The actions of working proteins are complex and have to be functionally synchronized, so continuous commands will be needed by all the amino acids in the protein so they can work together. The primary will also need continuous feedback to stay updated on the status of each position and the surrounding environment, just as humans are continuously updated with sensory data from all parts of the body even while we work.
If these many commands intended for many destinations were passed along by repetition at every position along the way, there could only be chaos. Clearly this communication system would not work for humans, and neither will it work for proteins.
For proteins to be functional, communication between the primary and every minderelle in the protein must be direct. For scientific, design, engineering, governing, administrative, and other protein types, direct and secure communications between any pair or grouping of minderelles will also be a requirement, just as this is required in human communications.
Addressing
Postal systems go back thousands of years. The effort to provide direct and secure communication between people separated by distance continues with today’s postal services, phones, computers, and electronics in the form of mail packages, conference calls, emails, texts, and social media. Humans have long understood the need for person to person connections, and more recently, person to group connections, over long distances.
So too would god have understood this same need for direct communications between the distant minderelles within a protein, and between the minderelles of different proteins connected by hydrogen bonds. In keeping with the methods of this chapter, mindesign mimics human design. Again, like minds using like reason solve like problems alike. So as per the methods of this chapter, we will accept human solutions to the problem of communicating over distances for the minderelles within a protein.
There are several necessary components to a telecommunication system. There is the means of transport – couriers, wires, radio waves, or the physical bodies of the minderelles making up a protein. Each message will include the address of both the sender and the recipient, thus making both delivery and replies possible. Each message will include the content of the message, and there must also be security, or confidence that the message was not altered between the sender and the recipient.
All of these requirements are generally met with any of today’s electronic communication methods. Each communication packet has the destination address, the source address, the message, and some level of security. Therefore, the hardware and software of today’s computer systems have, for the most part, solved the telecommunications problem. So how does telecommunication become possible for the minderelles in proteins?
We have the protein chain itself as a pathway for message transport. However, if messages are handled by the free thinking, mindcore portion of consciousness, we have lost security. This would be similar to a problem sometimes encountered in the telephone game, a single player that willfully disrupts the game by intentionally replacing the message with something not related.
For minderelles to have confidence that messages are not being altered en route, the messages will have to be handled by the fully determined mindshell portion of consciousness. Therefore, the mindshell will have programming similar to that inside the computers that handle today’s telecommunication packets.
However, one component of functional human mail systems is still missing for our protein’s telecommunication system – addresses. Addresses would not only make minderelle to minderelle communications possible, but would also solve another important problem we have not yet discussed. How does a protein prevent any minderelle in the protein from taking command and acting as the primary?
Thus, addresses will be used both to designate the primary of the protein, and to make secure communications between arbitrary groups of minderelles possible. The primary of a protein will always have address 0. In modern proteins, the primary of a protein would be designated by the ribosome when the protein is first assembled. To do this, the ribosome would externally assign the zero address to a designated position in each new protein during its assembly.
Once the zero minderelle takes primary status, this status will be maintained by the fully determined mindshells of the protein system. The mindshell of the primary will be responsible for sending out synchronization packets at the start of the mindshell’s timer cycle.
As these synchronization packets travel the length of the protein, each mindshell will increment or decrement the active address of the synchronization packet by one, depending upon the direction from the primary, thus designating or continuously confirming the permanent address of each minderelle in the protein.
Likely, minderelle addresses would have two parts. The first part would designate the minderelle’s count from the primary while the second part of the address would designate the number of hydrogen bond crossings for proteins joined together in beta sheets.
With addresses now assigned and maintained by the mindshells of the protein, we have all the infrastructure needed for direct and secure minderelle to minderelle messages, group messages, coordinated protein motion commands, and feedback. The mindcores of minderelles create messages for the intended recipients, then the mindshells of the protein handle the delivery logistics.
This is exactly the way it is with today’s computers. We hit the send button without a clue as to how anything works. All we know is that the message we wrote will somehow magically appear in the designated inboxes with our name as the sender. Engineers and programmers have taken care of all the details. In minderelles, the details are taken care of by the mindesign of mindshells.
Brain Synchronization
For humanons with a brain, the synchronization packets from the primary will be passed along through the full extent of the multiplexer in order to synchronize the switching of the multiplexer data paths so that the data type arriving at the primary will always remain in sync with the timer cycle of the primary. These primary synchronization packets will ensure that every multiplexer input line always has access to the primary during its corresponding time slot.
After the primary’s synchronization packets pass through the multiplexer, they will likely continue on until they have passed through all the portions of the brain where the primary’s input data is being processed. These primary timing signals will be used to keep all memory and sensory data sampling and processing synchronized with the multiplexer so that the data destined for each input line will be fully processed and properly timed for input into the primary during its corresponding time slot. The primary’s synchronization packets may pass through the command side of the brain as well and for similar reasons.
Because the human body, and other humanon bodies as well, have a nervous system that carries communications between the brain and distant parts of the body, it is possible that the primary’s synchronization packets will also be passed along throughout the body’s nervous system.
Operational Capabilities for Non-Primaries
We have been discussing the operational capabilities of conscious minds, but we have remained focused on humanon primaries. However, the vast majority of minderelles are not primaries. We will now focus on the minderelles that have non-primary roles within proteins that are inside cells.
We now recall that memory and the sensory fields are operational capabilities of the minderelle, so even non-primary minderelles will still have these same capabilities. But as non-primaries, these minderelles will have no incoming memory data arriving during the memory time slot of their timer cycle, and they will have no incoming sensory data arriving during their sensory data time slots.
For these non-primary minderelles, if there is no incoming data arriving during these time slots, then they will have access to their own memories and they will be free to populate their own sensory fields. Just as human artists and musicians use their imagination to create their original works, so will non-primary minderelles also be able to use their memories and imagination to populate their own sensory fields.
Once the content of a sensory field has been created, minderelles will be able share their creations with other minderelles within their communication network in the same way that we share photos across our communication networks. We have already covered this ability of minderelles to transfer sensory data – recall that the minderelles within the multiplexer were all transferring sensory data to the primary.
Thus, when design and engineering proteins are working on a new design for the flower of their new species, the minderelles of these engineering proteins will use their imagination and their communication capabilities to create, share, edit, and choose the design and color pattern of the petals, the fragrance of their new flower, and the taste of the nectar. With what we have covered so far, minderelles already have all the capabilities needed to share multi-sensory movies as well.
Interestingly, when design and engineering proteins work to create a beautiful flower for their newly modified plant species, their intention to create something beautiful reveals an understanding that their creation will be visually observed and appreciated by other conscious humanons.
To summarize, whenever memory and sensory field data are on a minderelle’s input line during the corresponding time slots, this incoming data will populate these operational fields and suppress the minderelle’s free access to these operational capabilities. When there is no incoming data during these time slots, the minderelles will have full access to their own operational capabilities. All of this will be governed by the mindshells of the minderelles.
Language Is Required for Design
Although the ability to transfer images and other sensory fields to one another is invaluable for engineering and technology purposes, it is also lacking. Scientists do not communicate sonar findings through visual images alone, they communicate their findings through a combination of visual images and language. Without language, many engineering feats would be next to impossible to understand, and other high level, cooperative efforts would also not be possible. The ability to communicate through an advanced language is necessary in all areas of society where cooperative efforts are required.
Cooperative efforts require many words that can express a wide array of concepts through a highly structured language. An advanced language will have clear definitions for its words, it will make use of all the sounds that can be produced by the vocal anatomy to maximize the rate of information transfer, it will have well defined parts of speech – nouns, verbs, adjectives, conjunctions, and so on – it will be easy to learn and use, so it will have intuitive spelling, construction, prefixes and suffixes, and grammar and punctuation rules. All of this will make rapid, unequivocal expressions of complex thoughts possible even for those with a more limited education.
Societal and world advancement is a function of advanced and universal communication capabilities. If great advances in technology, societal structures, and human affairs would not be possible for humans without advanced language capabilities, then neither should we expect such advances to be possible for the minderelles within proteins without advanced language capabilities. Reason is not scale dependent.
Without the exchange of information through an advanced language, the minderelles of engineering proteins could not be educated or kept abreast of advances in their field, and they could not properly submit their engineering proposals to others in their protein or protein network. And so it would be with proteins of every type. Accomplishments reveal capabilities.
For these reasons, we find that consciousness must include language as an innate operational capability.
Language Input
Some readers may have noticed that one of the inputs to the multiplexer was language.
The sound waves detected by the ears are processed by the brain and passed on to the primary where they will be heard by the mindcore during the audio time slot. Spoken words will be heard just as any other audio would be heard. Where speech is involved, though, the informational content of the words may have to be translated by the brain into a more universal language format that all mindshells can properly process for the mindcore.
The brain may be better suited for making this translation to a more universal language format because the brain may be designed for learning whatever language a child learns, and because the brain has free access to unsuppressed memory capabilities. Recall that the primary minderelle has incoming memory information arriving from the brain during the memory time slot, and this incoming memory data will suppress the ability of the primary to access its own memories.
The brain would also identify the speaker from the sound of the voice and pass this information along to the primary as well. Thus, the audio would enter the same as all audio data through the incoming shared bond during the audio time slot, while the translated informational content would enter the primary either through the hydrogen bonds of the primary, or possibly during a language time slot of the primary’s timer cycle.
When the mindcore decides on commands for the body, the mindshell will convert these commands into this same universal language format for export to the demultiplexer.
The Mindshell Tool Chest
Today’s computers provide us with database, spreadsheet, graphing, calculation, programming, and other capabilities. These tools have become indispensable in many areas of today’s science and engineering projects. Because these tools are created by way of programming code, and because the mindshell carries out all the programmable functions that make the life of minderelles fully functional and productive, it is likely that the mindshell will provide analogous computer-like capabilities to the mindcore.
As for the conscious interface experienced by the mindcore for these computer-like operations, it is likely the same interface we are familiar with. For a mindcore that is not a primary, it will be able to create the same rows and columns of a spreadsheet in its visual sensory field that we see on a computer screen. The mindcore will commit formulas to the cells and see their processing as the work of its own doing. And so it will be with the many other computer tools we are familiar with.
Because the mindcore has the programming capabilities of the mindshell supporting it, as well as the visual and audio sensory fields that are no different than our own, the minderelle’s computer capabilities are built-in rather than external.
The Identity of Humanon Minderelles
Clubs, schools, businesses, cities, nations, and other human groups will commonly keep their members informed of all the news and activities of the group. For their part, members of these human groups often identify as one with the groups to which they belong and feel a loyalty to their groups. Because the number of minderelles needed to keep humanons functional is extraordinarily high, it would be motivationally important to create a common identity for all the minderelles of a humanon.
Therefore, all the minderelles of a humanon will likely be kept informed of their humanon’s news and activities, will identify as one with their humanon, and will feel a loyalty to their humanon. Any of the minderelles from this humanon could later go on to become the primary of a human and this minderelle would still have all the memories of its previous experience with the earlier humanon.
While these memories would remain part of the minderelle’s personal memories, these memories would likely be inaccessible during the time it spends as a human primary, for these personal memories would be suppressed by incoming memories from the brain as discussed earlier. These memories would only be accessible should there be some disruption or lack of development with the incoming data from the brain during the memory time slot of the primary.